核酸認識TLRをターゲットとした治療戦略の可能性
| 村上 祐輔、山下 直美 | 武蔵野大学薬学部薬物療法学 |
| Yusuke Murakami, Naomi Yamashita | Research Institute of Pharmaceutical Sciences, Department of Pharmacology,Musashino University |
| 要 旨 | 肺や気管支は、呼吸や常在菌によって異物が恒常的に暴露される組織の一つである。したがって、病原体からの感染防御、有害物質による傷害、アレルギーや自己免疫疾患などの非感染性炎症、悪性新生物の排除など幅広い免疫応答が、生体の恒常性を維持するために機能している。著者らは、自然免疫受容体の応答が疾患の病態にどのように関わるのかに着目し研究してきた。近年、核酸認識Toll- Like Receptor (TLR)のリガンド応答機構を解析する過程で、マウスTLR3、TLR7、TLR9に対するモノクローナル抗体を独自に樹立した。 これらの抗体は、TLR応答を正または負に制御する機能を持つことを見出している。 本稿では、これらのユニークな機能を持つ抗体について紹介し、核酸認識TLRをターゲットとした新たな抗体医薬の可能性について述べる。 | |
|---|---|---|
| キーワード | 自然免疫、核酸認識Toll-Like Receptor(TLR)、抗体医薬 | |
| 『呼吸』eレポート 2(1) 1-8 ,2018 http://www.respiration.jp/erep/mokuji.php?y=2018&v=1 | ||
Ⅰ. 核酸認識TLR応答と疾患
核酸認識Toll- Like Receptor (TLR)は、ウイルスや細菌などの病原体由来の核酸を認識することに加えて、傷害を受けた自身の細胞、好中球から放出されるDNA(Neutrophil Extracellular Traps : NETs)、ミトコンドリアDNAなど自己由来の核酸をも認識して炎症性サイトカインやⅠ型インターフェロンの産生を誘導する1-3)。これら非感染性の炎症における自然免疫応答が、「自然炎症」という新たな概念として注目されている。有害物質による刺激、自己免疫疾患、肥満などによる生活習慣病、ガンなどの悪性新生物を発症すると、死細胞などから自己由来のリガンド(内因性リガンド)が健常時より多く発生する。内因性リガンドのうち核酸リガンドは、通常、核酸分解酵素(種々のDNaseやRNase)によって即座に分解されるため、核酸認識TLRが局在するライソソームに到達することはなく、免疫系の活性化も誘導されない。しかしながら、炎症病態においては、核酸に対する自己抗体やLL37のような核酸と複合体を形成するような蛋白質によって安定化し、酵素による分解をまぬがれる。そして、細胞内に取り込まれた核酸は、ライソソームに到達し、TLRを刺激して炎症を増悪することがよく知られている。例えば、TLR7やTLR9は、SLE(Systemic Lupus Erythematosus: 全身性エリテマトーデス)などの自己免疫疾患、慢性閉塞性肺疾患(COPD:chronic obstructive pulmonary disease)、アレルギー疾患に関わることが数多く報告されている4-9)。また、TLR8も、関節リウマチとの関連が報告されている10)。TLR応答が、病態を増悪させる例は、自己免疫疾患やアレルギー疾患だけではない。TLR3は、紫外線による皮膚傷害や放射線腸炎を増悪させることが近年、報告されている11, 12)。
以上のように、核酸認識TLRが内因性リガンドによって活性化し、炎症病態をさらに増悪させることは数多く示されており、有効な治療標的として特異的な核酸認識-TLR阻害剤の開発が求められている。
Ⅱ. 細胞表面に発現する核酸認識TLR
核酸認識TLRは、エンドソームまたはライソソームのような細胞小器官に発現して、細胞内に取り込んだリガンドを認識していることが知られていたため、抗体が直接、核酸認識TLRに結合してリガンド応答を制御するのは難しいと考えられてきた。一部の報告で、ヒトの細胞株に細胞表面TLR3が発現することが明らかとなっていたが、プライマリーな細胞で発現するのか、ヒトやマウスで検討されたことがなかった。そこで、著者らは、マウスTLR3、TLR7、TLR9に対するモノクローナル抗体をそれぞれ樹立して、マウスの骨髄から誘導したマクロファージの細胞表面を染色したところ、TLR3、TLR7は、細胞表面に発現していることが示された13, 14)(図1A)。さらにTLR9は、脾臓の樹状細胞(cDC:conventinal Dendritic cell)および形質
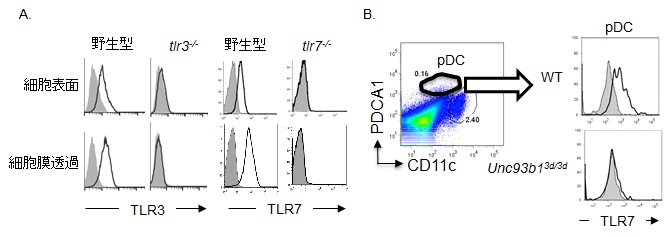
A.マウス骨髄から誘導されたマクロファージのTLR3およびTLR7の発現を解析.
B.マウス脾臓pDCにおける細胞表面TLR7のUnc93B1依存性.
灰色:抗体なし, 黒線:抗体あり
細胞様樹状細胞(pDC:plasmacytoid Dendritic cell)で、細胞表面に発現していることを報告している15, 16)。これらのTLRを介するリガンド応答には、Unc-93 homolog B1(Unc93B1)によるエンドライソソームへの細胞内輸送が重要であると報告されている17)。そこで、細胞表面への移行にもUnc93B1が必要であるかを検討した。著者らは、野生型と、Unc93B1の機能不全変異体マウスであるUnc93B13d/3d マウスの脾臓で、pDCの細胞表面TLR7発現を比較した。その結果、細胞表面TLR7の発現には、Unc93B1が必要なことを示した(図1B)。
筆者らの検討で、細胞表面の核酸認識TLRが、抗体の標的になりうることを示すことができたが、免疫応答にどのような役割を持つのか、どのような細胞内移行が起こっているのか、未だ不明な点があり、さらに詳細な解析が期待される。
Ⅲ. TLR-抗体複合体の安定的な
細胞内取り込み
細胞内取り込み
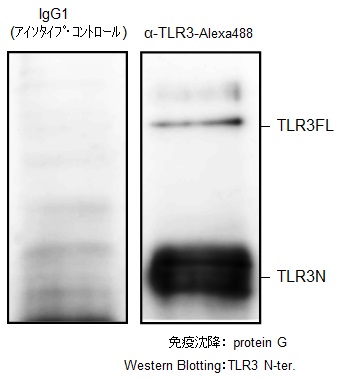
核酸認識TLRに対する抗体が、細胞表面のTLRに結合したのちにどのような挙動を示すのかを検証した。まず、蛍光色素Alexa488を付加したTLR3抗体(α-TLR3-Alexa488)をマウス由来J774.1細胞に添加し37℃または4℃で培養し検討した(図2A)。4℃での反応は、Alexa488の蛍光を消失させる抗体によって蛍光が消えたため、α-TLR3-Alexa488は細胞表面のTLR3には結合するものの、細胞内への取り込みは起こらないことが示唆された。また、37℃での培養ではα-TLR3-Alexa488による蛍光の消失が起こらないため、抗体は細胞内に取り込まれたことが示唆された。さらに、α-TLR7-Alexa488を、TLR7とUnc93B1を強制発現させたマウス線維芽細胞株(NIH-3T3)と培養して4時間後の細胞を共焦点レーザー顕微鏡で観察した。α-TLR7-Alexa488の蛍光が、核の周囲、細胞質に観察され、抗体が細胞内に取り込まれたことが示された(図2B左)。TLR7と同時にライソソーム(LAMP-1陽性)を染色すると、TLR7とライソソームが共局在することが観察された(図2B右、矢頭)。続いて、取り込まれた抗体がTLRと結合して、細胞内で安定した複合体を形成できるのかを検討した。J774.1細胞にα-TLR3-Alexa488を加えて37℃で一晩培養した。フローサイトメーターで細胞の蛍光を確認後、それを溶解して、プロテインGビーズで抗体ごとTLR3を沈降できるのかを検討した。その結果、TLR3は、TLR3抗体を加えたカラムで特異的に検出された(図3)。以上の結果より示すことができた。
Ⅳ. TLR3応答を正に制御する
モノクローナル抗体
モノクローナル抗体
TLR3は、二重鎖RNA(dsRNA)や合成リガンドであるpolyI:C(ポリイノシン酸:ポリシチジン酸)を認識して、炎症性サイトカインやⅠ型インターフェロン産生を誘導する。通常、われわれの体内ではdsRNAがうまれることがないためウイルス由来のdsRNAがTLR3の主なリガンドとして考えられてきた。しかしながら、TLR3は日焼けや放射線腸炎を増悪することが報告され注目されている11, 12)。すなわち、ある条件下でうまれるRNAは、たとえ一本鎖であってもTLR3のリガンドになり得ることが示されたことで、新たな展開が期待されている。
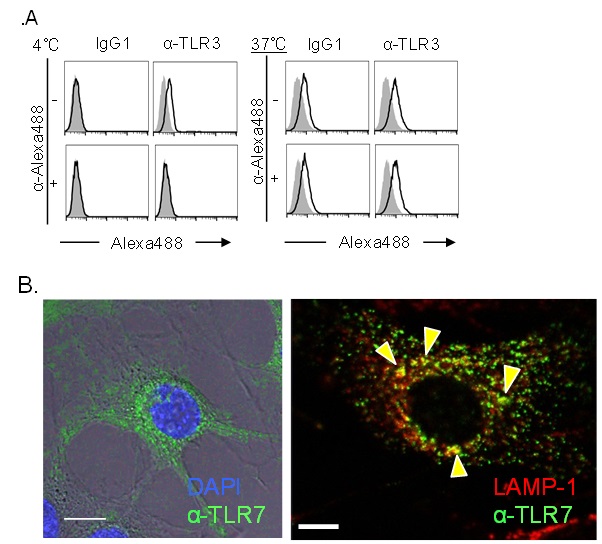
A.J774.1細胞におけるTLR3抗体の取り込み. 灰色:抗体なし, 黒線:抗体あり.
B.TLR7抗体のNIH-3T3細胞への取り込み(左図).
ライソソーム(LAMP-1)とTLR7の共局在(右図 黄矢頭).
ほかのTLRシグナルでは見られないTLR3の特徴として、クロスプレゼンテーションを促進してウイルス感染や腫瘍免疫を促進する機能が知られている18-20)。TLR3応答のみが他のTLR応答と比較して、どのようにクロスプレゼンテーションを活性化するのか、未だに詳しく分かっていない。さらにpolyI:Cは、TLR3だけでなく、MDA5(Melanoma Differentiation-Associated protein 5)などの細胞質受容体も刺激して強い免疫応答を誘導するため治療薬への応用が難しい。そこで、特異的にTLR3を活性化する新たな方法が求められている。
著者らは、TLR3のリガンド認識機構を解明する過程で、新たなモノクローナル抗体(クローン名:PaT3)を樹立した14)。骨髄細胞からマクロファージを誘導し、PaT3とpolyI:Cを併用して刺激するとRANTES/CCL5の産生を特異的に増強した(図4A)。さらに、in vivoでの増強効果を検証するために、マウスの腹腔内に抗体(200μg/head)を投与後、24時間後にpolyI:C 50μg/mlを投与して経時的に血清中のIL-12p40を測定した。その結果、アイソタイプコントロールである
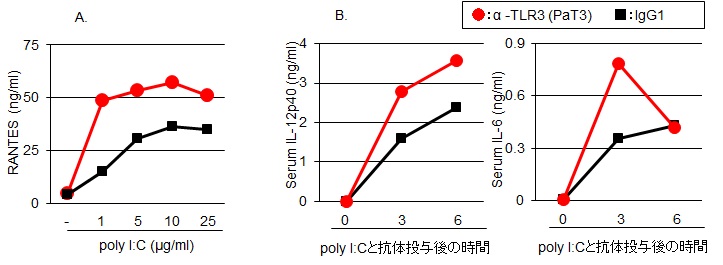
A.骨髄細胞由来マクロファージをPaT3またはIgG1とpolyI:Cを併用して刺激した.
B. in vivoでのTLR3応答増強効果.
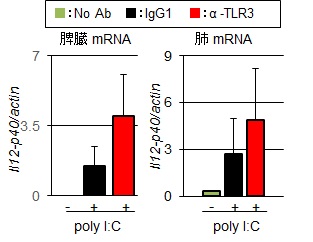
IgG1とpolyI:Cを併用したマウスと比較して、IL-12p40産生が増強された(図4B)。さらに, 投与後6時間の脾臓と肺でのIL-12p40のmRNA発現量を測定した。PaT3とpolyI:Cを投与したマウスでは、コントロールと比較して脾臓および肺でのIL-12p40の発現は有意に(p<0.05)高かった(図5)。今後、肺局所でのウイルス感染や腫瘍の退縮効果を明らかにし、TLR3抗体を用いた新たな免疫活性法の有効性を検証したいと考えている。さらに、内因性リガンドを含めたpolyI:Cに代わる新たなTLR3リガンドを明らかにすることで、TLR3抗体投与による新たな免疫療法を確立できると考えている。
Ⅴ. TLR7 およびTLR9応答を負に制御する
モノクローナル抗体と病態制御
モノクローナル抗体と病態制御
TLR7は、病原体由来の一本鎖RNAやグアノシン誘導体を認識するが、一方で、自己免疫疾患モデル動物やSLE患者由来の検体の解析から、内因性RNAリガンドを認識して様々な病態に関わることが示唆されている4-9)。アレルギー喘息では、TLR7リガンド刺激が病態を抑制するという報告と増悪するという相反する報告がある。リガンド認識について最近の研究で、核酸由来のグアノシンやデオキシグアノシンを認識することが報告され、より詳細な内因性リガンド認識の分子メカニズムが明らかにされつつある21, 22)。
著者らは、独自に樹立したマウスTLR7に対するモノクローナル抗体(クローン名:A94B10)を用いて、マウス脾臓および骨髄から誘導した免疫細胞の解析により、TLR7が、マクロファージ、cDC、pDC、B細胞の細胞表面に発現することを明らかにしている13)。また、A94B10は、in vitroでマクロファージ、cDC、pDCのTLR7応答を効果的に阻害した(図6)。さらに、in vivoでの阻害効果を検証するために、TLR7依存的な致死性炎症を自然発症するUnc93B1-D34A変異体ノックインマウス23)に、この抗体を投与して治療効果を検証した。このマウスは、自己抗体の産生や血小板減少が起こり、肝臓、脾臓、腎臓など全身に炎症を起こして30週齢までに約50%が死亡する(図7)。筆者らは、このマウスにA94B10を投与して治療効果を検証した。その結果、肝臓における免疫細胞の浸潤、線維化などの炎症を著しく抑制した(図8)。さらに、血小板数の減少、脾臓の肥大化などを回復させた。著者らは、TLR7応答が病態に関わるヒト疾患を探索中であり、様々な疾患モデルにA94B10を投与して治療効果の検証を進めている。
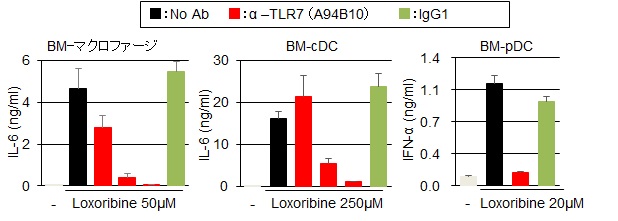
マクロファージ, cDC, pDCにおけるTLR7応答阻害効果. TLR7抗体群(赤)は, 左から0.001, 0.1, 10μg/mlの抗体濃度. pDCでは, 10μg/ml. コントロール抗体(IgG1)群は, 10μg/ml
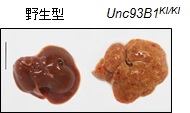
TLR7依存的な致死性炎症を発症している。 Bar : 1 cm
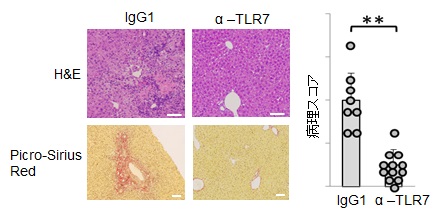
Bar : 100μm. ** : p<0.001
TLR9は、DNAを認識する受容体として感染症、自己免疫疾患、COPDの病態に関わることが報告されている1-6)。TLR9の膜貫通ドメインをTLR4の膜貫通ドメインに置換したノックインマウスでは、TLR9の局在がライソソームから細胞表面に移行して、細胞外のDNAを認識するため致死性の炎症シグナルを誘導することが報告されていた24)。著者らは、マウスTLR9のモノクローナル抗体を4クローン樹立し報告している15, 16)。
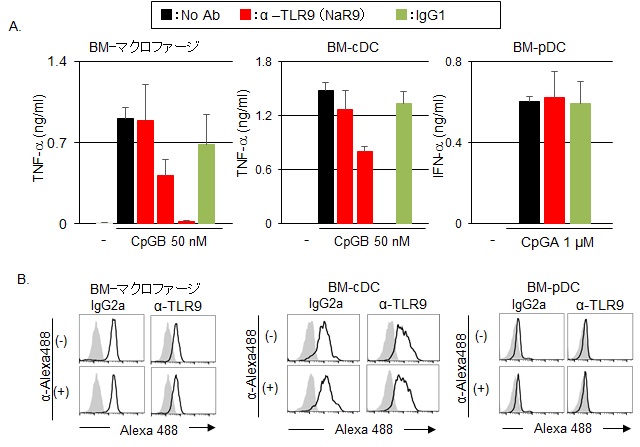
A.マクロファージでは, TLR9抗体を, 左から0.1, 1, 10μg/mlの抗体濃度. コントロール抗体群は, 10μg/mlで用いた. cDCでは, TLR9抗体を, 左から0.3, 3, 30μg/mlの抗体濃度. コントロール抗体群は, 30μg/mlで用いた. pDCでは, 抗体を30μg/mで用いた.
B.TLR9抗体の細胞への取り込み. 灰色:抗体なし, 黒線:抗体あり.
その中で、クローン名:NaR9は、骨髄から誘導したマクロファージおよびcDCで特異的にTLR9リガンド(CpGB)によるTLR9応答を阻害した。一方、pDCのTLR9応答は阻害しなかった(図9A)。この原因を解明するために、TLR9抗体の細胞への取り込みを検証した。TLR9抗体にAlexa488を付加した α-TLR9-Alexa488を、骨髄から誘導したマクロファージ、cDC、pDCに加え 37℃で一晩培養した。細胞の表面に結合した抗体の蛍光を除くために、蛍光を消失させるAlexa488抗体で細胞を処理した後、フローサイトメーターで蛍光を測定した。その結果、マクロファージ、cDCでは、TLR9抗体とコントロール抗体は共に細胞内に取り込まれたが、pDCでは細胞への取り込みを確認できなかった(図9B)。以上により、pDCでTLR9応答を阻害できない原因は、抗体が細胞内に取り込まれないためであることが示唆された。
続いて in vivoでの阻害効果を検証するために、TLR9リガンド(CpGB)とD-(+)-ガラクトサミンの併用で致死的な肝臓の炎症を誘導する実験系を用いた。その結果、コントロール群(PBS群、IgG2a群)では約90%が刺激から一日以内に死亡したが、TLR9抗体投与群では死亡率が大きく抑えられ、80%が刺激から5日目以降も生存した(図10左)。この実験系は、肝臓でのTNF-αの産生が致死的な肝細胞死を誘導することが知られている。そこで、本実験のマウスから経時的に採血を行い、血清中のTNF-α濃度を測定した。その結果、TLR9抗体(NaR9)投与群では有意にTNF-α産生が抑制された(p<0.05 図10右)。
VI. 今後の展望
核酸認識TLRは、病原体由来の核酸を認識する受容体として発見され、病原体の排除や宿主自身への過剰な炎症反応を誘導することが知られている。また、多くの報告で自己免疫疾患などとの関連が述べられている。
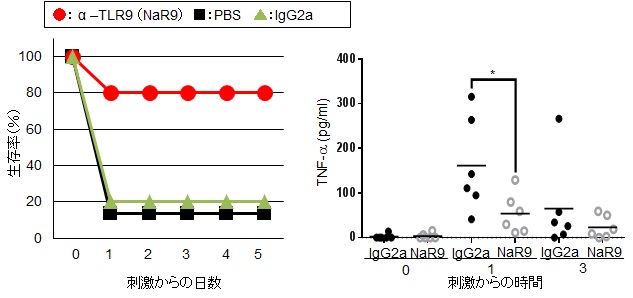
* : p<0.05
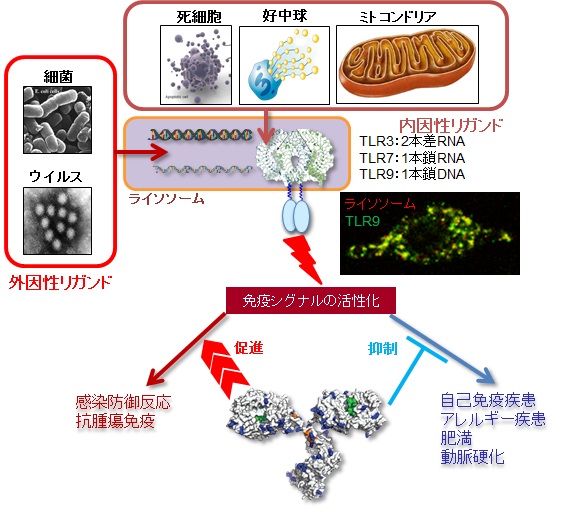
筆者らは、マウスTLR3、TLR7、TLR9に対するモノクローナル抗体を用いて核酸認識TLRのリガンド応答を正または負に制御することを報告してきた12, 13, 15)。今後、ヒトTLRを正または負に制御する抗体を用いて、新規のヒト疾患治療薬への応用を目指して行きたい(図11)。
TLR研究の進展は、ヒト難治性疾患とTLR応答の関連を解明しつつある。今後さらに、核酸認識TLRを標的とした核酸や低分子化合物の開発競争も増すことが予想され、新たな展開が期待される。
1) Sisirak, V. et al. Digestion of Chromatin in Apoptotic Cell Microparticles Prevents Autoimmunity. Cell 166:88.
2) Garcia-Martinez, I. et al. Hepatocyte mitochondrial DNA drives nonalcoholic steatohepatitis by activation of TLR9. J Clin Invest 126:859.
3) Okabe, Y. et al. Toll-like receptor-independent gene induction program activated by mammalian DNA escaped from apoptotic DNA degradation. J Exp Med 202:1333. 2005.
4) Christensen, S. R. et al. Toll-like receptor 7 and TLR9 dictate autoantibody specificity and have opposing inflammatory and regulatory roles in a murine model of lupus. Immunity 25:417.
5) Zhu, F. G. et al. A novel antagonist of Toll-like receptors 7, 8 and 9 suppresses lupus disease-associated parameters in NZBW/F1 mice. Autoimmunity 46:419.
6) Hung, T. et al. The Ro60 autoantigen binds endogenous retroelements and regulates inflammatory gene expression. Science 350:455.
7) Foronjy, F. R. et al. TLR9 expression is required for the development of cigarette smoke-induced emphysema in mice. Am J Physiol Lung Cell Mol Physiol 311: L154–L166.
8) Deane, J. A. et al. Control of toll-like receptor 7 expression is essential to restrict autoimmunity and dendritic cell proliferation. Immunity 27:801.
9) Adner, M. et al. Toll-like receptor (TLR) 7 decreases and TLR9 increases the airway responses in mice with established allergic inflammation. Eur J Pharmacol 718:544-551.
10) Guiducci, C. et al. RNA recognition by human TLR8 can lead to autoimmune inflammation. J Exp Med 210:2903.
11) Belnald, J. J. et al. Ultraviolet radiation damages self noncoding RNA and is detected by TLR3. Nat Med. 18:1286-1291.
12) Takemura, N. et al. Blockade of TLR3 protects mice from lethal radiation-induced gastrointestinal syndrome. Nat Commun. 5:3492.
13) Kanno, A. et al. Targeting cell surface TLR7 for therapeutic intervention in autoimmune diseases. Nat Commun 6:6119.
14) Murakami, Y. et al. Roles of the cleaved N-terminal TLR3 fragment and cell surface TLR3 in double-stranded RNA sensing. J Immunol 193:5208
15) Onji, M. et al. An essential role for the N-terminal fragment of Toll-like receptor 9 in DNA sensing. Nat Commun 4:1949.
16) Murakami, Y. and Fukui, R. et al. The protective effect of the anti-Toll-like receptor 9 antibody against acute cytokine storm caused by immunostimulatory DNA. Sci Rep 7:44042.
17) Kim, Y-M. et al. UNC93B1 delivers nucleotide-sensing toll-like receptors to endolysosomes. Nature 452:234-239.
18) Schulz, O. et al. Toll-like receptor 3 promotes cross-priming to virus-infected cells. Nature 433:887-892.
19) Levine, A. S. & Levy, H. B. Phase I-II trials of poly IC stabilized with poly-Llysine. Cancer Treat. Rep. 62:1907–1912.
20) Seya, T. et al. Role of Toll-like receptors and their adaptors in adjuvant immunotherapy for cancer. Anticancer Res. 23:4369–4376.
21) Zhang, Z. et al. Structural Analysis Reveals that Toll-like Receptor 7 Is a Dual Receptor for Guanosine and Single-Stranded RNA. Immunity 45:737.
22) Shibata, T. et al. Guanosine and its modified derivatives are endogenous ligands for TLR7. Int Immunol 28:211.
23) Fukui, R. et al. Unc93B1 restricts systemic lethal inflammation by orchestrating Toll-like receptor 7 and 9 trafficking. Immunity 35:69.
24) Mouchess, L. M. et al. Transmemrane Mutations in Toll-like Receptor 9 Bypass the Requirement for Ectodomain Proteolysis and Induce Fatal Inflammation. Immunity 35:721-732.